Abstractions/Abstract concept generator
[[Image:Tursiops truncatus 01.jpg|thumb|right|250px|A bottlenose dolphin surfs the wake of a research boat on the Banana River. Credit: NASA.{{tlx|abagar lus
"They talk about an "abstract concept generator" [a generator or generative] which produces “some kind of abstract object” [that] represents the maximal content of a whole set of discourse deriving from this concept."[1]
Abstractions
editDef.
- the "act of focusing on one characteristic of an object rather than the object as a whole group of characteristics;"[2]
- "the act of separating said qualities from the object or ideas",[3]
- the "act of comparing commonality between distinct objects and organizing using those similarities;"[2]
- "the act of generalizing characteristics; the product of said generalization",[3]
is called abstraction.
Conceptualizations
editAs "the body (of individuals and of a comparative series of adults of diffèrent species) grows larger, the brain grows larger in some regular fashion."[4]
Generators
editDef. that which is
- an "apparatus [forming] vapour or gas [from] a liquid or solid by means of heat or chemical process, as a steam boiler, gas retort, or vessel for generating carbonic acid gas, etc.",[5]
- a "principal sound or sounds by which others are produced; the fundamental note or root of the common chord; -- see also generating tone",[5]
- an "element of a group that is used in the presentation of the group: one of the elements"[6] "from which the others can be inferred with the given relators",[7]
- one "of the lines of a ruled surface; more generally, an element of some family of linear spaces",[8]
- a "subordinate piece of code which, given some initial parameters will generate multiple output values on request",[7]
- a "piece of apparatus, equipment, etc, to convert or change energy from one form to another",[9] or
- "a machine that converts mechanical energy into electrical energy"[10]
is called a generator.
Reifications
editDef. the "consideration of an abstract thing as if it were concrete, or of an inanimate object as if it were living"[11] is called reification.
Theory of abstract concept generation
editHere's a theoretical definition:
Def. a sufficiently large neural net to convert one or more sensory concepts into an abstract concept is called an abstract concept generator.
Earth
editOn Earth past and present have been and are a number of species whose brain or central nervous system size, mass, or neuronal complexity at least in adults is necessary and sufficient to allow most individuals to think and conceive in the abstract. This allows them to make or utilize tools where possible or exhibit certain intuitive skills which are ear marks of the possession of an abstract concept generator.
Cetaceans
editSignificant "evidence [exists] of positive selection on the [abnormal spindle-like microcephaly associated] ASPM gene during cetacean evolution, especially for the Odontoceti and Delphinoidea lineages."[12]
"Primates and cetaceans are the most remarkable examples of such massive brain size expansion, especially of the cerebral cortex—the brain area associated with higher cognitive functions [3]."[12]
The "evolution of larger brain sizes in these groups has been driven by selection for life in increasingly complex social environments and the need for highly developed cognitive abilities [4,5]. Accordingly, the primate brain, and especially the human brain, has been recognized as one of the most striking evolutionary adaptations [6,7]."[12]
"Cetaceans diverged from terrestrial artiodactyls approximately 56 – 53 million years ago (Ma) [8]".[12]
"Early cetaceans (called archaeocetes) diversified through amphibious stages to become fully aquatic by 40 Ma [9]. Extant cetaceans, consisting of highly diversified species subdivided into two suborders, Mysticeti (large rorqual and baleen whales) and Odontoceti (dolphins, porpoises and toothed whales), evolved from archaeocetes at about 34 Ma and dispersed into the world’s oceans and estuaries and even some rivers [10,11]."[12]
Paleontological "and neuroanatomical data have confirmed that modern cetacean brains are among the largest across all mammals both in absolute and relative size (expressed relative to body size as encephalization level or quotient, EQ [1]). Almost all odontocetes (toothed whales) have above average levels of encephalization compared with relative terrestrial mammals. In particular, some odontocetes possess EQs in the range of 4 – 5, second only to modern humans (EQ ≈ 7) and significantly higher than any of the modern nonhuman anthropoid primates (highest EQ ≈ 3.3) [15,16]."[12]
Sperm whales
edit[[Image:Sperm whale Tim Cole NMFS crop.jpg|thumb|right|250px|The image is of a sperm whale. Credit: Tim Cole, NMFS (NOAA).{{free media}}]] The image at the right is an aerial view of an adult sperm whale Physeter macrocephalus. "Sperm whales have the largest brain of any animal (on average 17 pounds (7.8 kg) in mature males)".[13] These sentient whales have the largest brains that have ever existed on Earth.
"Absolute size is the most general of all brain properties [...], and ranges in mammals from brains of small bats and insectivores (weighing less than 0.1 g) to those of large cetaceans (up to 9000 g)."[14]
Omaru whales
edit
"Omura whales have been reported at sizes of up to 124.5 feet."[15]
"This gentle giant [shown in the center frontal profile image], is shy and subtle."[15]
The holotype is an 11.03 m (36.2 ft) adult female, NSMT-M32505 (National Science Museum, Tokyo), which stranded at Tsunoshima (34°21'03"N 130°53'09"E) in the southern Sea of Japan on 11 September 1998. It includes a complete skeleton, both complete rows of baleen plates, and frozen pieces of muscle, blubber, and kidney collected by T. K. Yamada, M. Oishi, T. Kuramochi, E. Jibiki, and S. Fujioka. The type locality is the Sea of Japan, which may not be representative of the species’ typical range. The paratypes include the eight specimens (five females and three males), NRIFSF1-8 (National Research Institute of Far Seas Fisheries, Fisheries Research Agency, Shizuoka), collected by Japanese research vessels in the Indo-Pacific in the late 1970s. The longest baleen plate (NRIFSF6 includes 18 more baleen plates), an earplug, and a piece of the sixth thoracic vertebra with associated epiphysis were collected from each individual.[16][17]
Blue whales
edit
The brain mass of one blue whale adult is 5,678 gms.[18] Although another adult was measured as about 6800 gms.[19]
"The blue whale, our largest known animal is about 98.5 feet long."[15]
Killer whales
edit

The mean brain mass of Orcinus orca for six adults is 6,368 gms.[18]
Killer whales have the second-heaviest brains among marine mammals[20] (after sperm whales, which have the largest brain of any animal).
The killer whale's use of dialects and the passing of other learned behaviors from generation to generation have been described as a form of animal culture.[21]
Beaked whales
edit"Beaked whales — a family [Ziphiidae] of 22 cetacean species [in six genera (Berardius, Hyperoodon, Indopacetus, Mesoplodon, Tasmacetus, and Ziphius)] characterized by dolphinlike noses and missile-shaped bodies — are some of the most elusive animals on Earth. They dive deeper and longer than any other marine mammal and spend an estimated 92 percent of their lives far beneath the ocean surface. One species, the True's beaked whale, is so rare that only a handful of people have ever seen it alive."[22]
"Imagine, these are animals the size of elephants that we just can't find. They're a mystery."[23]
"We don’t know how large the populations of True's beaked whale or any other species are. The populations could decline and we would never know."[23]
"Since the species was identified by Smithsonian scientist Frederick William True in 1913, dead True's beaked whales (Mesoplodon mirus) have been found washed up on beaches across the North Atlantic and South Pacific."[22]
"Both male and female True's beaked whales are remarkably well-adapted to their deep-sea lifestyle. Their rotund bodies are torpedo shaped, and their sides bear “pockets” into which they can tuck their flippers, making them even more hydrodynamic. They have never been tagged, so scientists don't know how deep they can swim. But if the habits of other beaked whale species are anything to go by, they are certainly champion divers; in 2014, scientists at the Scripps Institute of Oceanography reported that a Cuvier's beaked whale journeyed 9,816 feet below the surface, breaking a marine mammal record. The animals can spend hours underwater on a single breath of air."[22]
In the second image down on the right is a fossil skull of Mystacodon, a 36-million-year-old whale found in Peru, is an early relative of today’s baleen whales.
"Based on an articulated cetacean skeleton [close to 4 meters long] from the early late Eocene (Priabonian, around 36.4 million years ago) of the Pisco Basin, Peru, we describe a new archaic tooth-bearing mysticete, Mystacodon selenensis gen. et sp. nov. Being the geologically oldest neocete (crown group cetacean) and the earliest mysticete to branch off described so far, the new taxon is interpreted as morphologically intermediate between basilosaurids and later toothed mysticetes, providing thus crucial information about the anatomy of the skull, forelimb, and innominate at these critical initial stages of mysticete evolution."[24]
"The genus Berardius has two species, Baird’s beaked whale Berardius bairdii, found in the North Pacific and adjacent waters, and Arnoux’s beaked whale B. arnuxii, found in the Southern Ocean3."[25]
"The whales differed from all of their congeners by having the following unique characters: a substantially smaller body size of physically mature individuals, proportionately shorter beak, and darker body colour. Thus, we conclude that the whales are a third Berardius species [Berardius minimus]."[25]
The "two species [Berardius bairdii and Berardius arnuxii]16 are genetically distinct and independent."[25]
"At least in adult and subadult individuals of B. minimus, cookie-cutter shark bites are fairly conspicuous, but not to the extent as usually seen in some other species such as Ziphius cavirostris, Mesoplodon densirostris, and/or Balaenoptera borealis."[25]
"If “karasu” exists as a third type, it could be a species that is not yet recognised or a Mesoplodon species found in Hokkaido (either M. stejnegeri or M. carlhubbsi). Recognition of these Mesoplodon species around Hokkaido is rather recent; the earliest M. stejnegeri specimen was collected in 198533, and the earliest M. carlhubbsi in 200434."[25]
Extinct genera of the subfamilia Berardiinae include Africanacetus, Archaeoziphius and Microberardius.[26]
Genus Hyperoodon has two species: H. ampullatus and H. planifrons.[27]
Genus Indopacetus has one species: I. pacificus.[28]
Genus Mesoplodon has 12 species: M. bidens, M. bowdoini, M. carlhubbsi, M. densirostris, M. europaeus, M. ginkgodens, M. grayi, M. hectori, M. layardii, M. mirus, M. peruvianus, and M. stejnegeri.[29]
Genus Mesoplodon has the additional species: M. hotaula[30], M. perrini[31], †M. slangkopi[32], M. traversii[33], and †M. tumidirostris[34].
Dolphins
editLike humans and dogs the brain of the toothed whales sits behind the eyes. The melon often seen on the front of the head of Tursiops truncatus (the common bottlenose dolphin) is not where its brain is located. The melon is used in echo-location.
Bottlenose dolphins
edit"Dolphins evolved from relatively small-brained animals like cows and hippos into this large-brained, highly specialized aquatic organism, [...] This is one of the first comprehensive studies to look at rates of molecular evolution in dolphins."[35]
"Because dolphins have also evolved large brains, it gives us an example of the independent evolution of big, complex brains to compare to the evolution of the human brain [...] By doing this, you can find out what is necessary for a big brain."[35]
Both humans and dolphins have mutations in the microcephalin gene.[36]
At the top of this resource is an image of Tursiops truncatus, the bottle-nosed dolphin, which in an average adult has a brain weight of about 2200 gms. One Tursiops truncatus adult female had a brain weight of 1,609 gm.[18] Several males averaged 1638 gms, but the ranges are 1,112-1,609 gms (females) and 954-1,910 gms (males) with an overall species average of 1,489 gms.[18]
Grand dolphins
edit
The image at the right is of a grand dolphin, Tursiops aduncus, of the Indian Ocean. Here it is photographed in the Port River, Adelaide, Australia.
The brain size of this dolphin may be similar to that of the bottle-nosed dolphin.
Proboscideans
edit
| |||||||||||||||||||||||||||||||||||||||||||||
| Proboscidea phylogeny is based on Shoshani 1998.[37] |




Over 185 extinct members and three major evolutionary (adaptive) radiations of the order Proboscidea have been recorded.[38]
The earliest proboscids, the African Eritherium and Phosphatherium of the late Paleocene, heralded the first radiation.[39]
The Eocene included Numidotherium, Moeritherium, and Barytherium from Africa. These animals were relatively small and aquatic, where, later on, genera such as Phiomia and Palaeomastodon arose; the latter likely inhabited forests and open woodlands.
Proboscidean diversity declined during the Oligocene.[40] One notable species of this epoch was Eritreum melakeghebrekristosi of the Horn of Africa, which may have been an ancestor to several later species.[41]
The beginning of the Miocene saw the second diversification, with the appearance of the Deinotheriidae deinotheres and the Mammutidae, the mammutids. The former were related to Barytherium and lived in Africa and Eurasia,[42] while the latter may have descended from Eritreum[41] and spread to North America.[42]
The second radiation was represented by the emergence of the gomphotheres in the Miocene,[42] which likely evolved from Eritreum[41] and originated in Africa, spreading to every continent except Australia and Antarctica. Members of this group included Gomphotherium and Platybelodon.[42]
The third radiation started in the late Miocene and led to the arrival of the elephantids, which descended from, and slowly replaced, the gomphotheres.[43] The African Primelephas gomphotheroides gave rise to Loxodonta, Mammuthus, and Elephas; Loxodonta branched off earliest around the Miocene and Pliocene boundary while Mammuthus and Elephas diverged later during the early Pliocene; Loxodonta remained in Africa while Mammuthus and Elephas spread to Eurasia, and the former reached North America; the Stegodontidae (stegodontids), another proboscidean group descended from gomphotheres, spread throughout Asia, including the Indian subcontinent, China, southeast Asia, and Japan; mammutids continued to evolve into new species, such as the American mastodon.[44]
Proboscideans experienced several evolutionary trends, such as an increase in size, which led to many giant species that stood up to 5 m (16 ft) tall.[45] As with other megaherbivores, including the extinct sauropod dinosaurs, the large size of elephants likely developed to allow them to survive on vegetation with low nutritional value.[46] Their limbs grew longer and the feet shorter and broader.[37] The feet were originally plantigrade and developed into a digitigrade stance with cushion pads and the sesamoid bone providing support.[47]
Early proboscideans developed longer mandibles and smaller craniums while more derived ones developed shorter mandibles, which shifted the head's center of gravity. The skull grew larger, especially the cranium, while the neck shortened to provide better support for the skull. The increase in size led to the development and elongation of the mobile trunk to provide reach; the number of premolars, incisors and canines decreased.[37] The cheek teeth (molars and premolars) became larger and more specialized, especially after elephants started to switch from C3 carbon fixation plants to C4 carbon fixation grasses, which caused their teeth to undergo a three-fold increase in teeth height as well as substantial multiplication of lamellae after about five million years ago; but only in the last million years or so did they return to a diet mainly consisting of C3 trees and shrubs.[48][49] The upper second incisors grew into tusks, which varied in shape from straight, to curved (either upward or downward), to spiralled, depending on the species, while some proboscideans developed tusks from their lower incisors.[37] Elephants retain certain features from their aquatic ancestry, such as their middle ear anatomy.[50]
Deinotherium giganteum
edit



Two adults of Deinotherium giganteum around 3.63–4 metres (11.9–13.1 ft) tall and weighing 8.8–12 tonnes (8.7–11.8 long tons; 9.7–13.2 short tons) are smaller than a 45-year-old male of Deinotherium "thraceiensis", at 4.01 metres (13.2 ft) tall and 13.2 tonnes (13.0 long tons; 14.6 short tons).[51]
Deinotherium is distinguished from its predecessor Prodeinotherium by its much greater size, greater crown dimensions, and reduced development of posterior cingula ornamentation in the second and third molar.[52]
Early Pleistocene species of Deinotherium might have also fallen prey to the sabertooth Homotherium, particularly adolescents and calves.[53]
A tooth of a deinothere found on the island of Crete, in shallow marine sediments of the Miocene[54] suggests that Crete was closer or connected to the mainland during the Messinian salinity crisis.
Mammut americanum
edit
Stegodonts
edit
Stegodonts were present from 11.6 mya to late Pleistocene, with unconfirmed records of regional survival until 4,100 years ago. Fossils are found in Asian and African strata dating from the late Miocene. They lived in large parts of East and Central Africa during the Pleistocene.[55][56]
Elephants
editElephants are in the family Elephantidae, the sole remaining family within the order Proboscidea which belongs to the superorder Afrotheria, where their closest extant relatives are the sirenians (dugongs and manatees) and the hyraxes, with which they share the clade Paenungulata within the superorder Afrotheria.[57]
Elephants and sirenians are further grouped in the clade Tethytheria.[58]
Three species of living elephants are recognised; the African bush elephant (Loxodonta africana) and African forest elephant (Loxodonta cyclotis) of sub-Saharan Africa, and the Asian elephant (Elephas maximus) of South and Southeast Asia.[59]
African elephants have larger ears, a concave back, more wrinkled skin, a sloping abdomen, and two finger-like extensions at the tip of the trunk.
Asian elephants have smaller ears, a convex or level back, smoother skin, a horizontal abdomen that occasionally sags in the middle and one extension at the tip of the trunk. The looped ridges on the molars are narrower in the Asian elephant while those of the African are more diamond-shaped. The Asian elephant also has dorsal bumps on its head and some patches of depigmentation on its skin.[60]
Genetics
editDNA studies suggest Mammuthus is more closely related to Loxodonta.[61][62]
Others point to Elephas.[58]
However, analysis of the complete mitochondrial genome profile of the woolly mammoth (sequenced in 2005) supports Mammuthus being more closely related to Elephas.[63][64][65][66]
Morphological evidence supports Mammuthus and Elephas as sister taxa while comparisons of protein albumin and collagen have concluded that all three genera are equally related to each other.[67]
Some scientists believe a cloned mammoth embryo could one day be implanted in an Asian elephant's womb.[68]
Palaeoloxodon
edit
The "recovery of full mitochondrial genomes from four and partial nuclear genomes from two P. antiquus fossils [...] collected at two sites in Germany, Neumark-Nord and Weimar-Ehringsdorf, and likely date to interglacial periods ~120 [MIS 5e] and ~244 [MIS 7] thousand years ago, respectively [white dots on the Eurasian fossil distribution map second down on the right] suggest that P. antiquus was a close relative of extant African forest elephants (Loxodonta cyclotis)."[69]
The "European straight-tusked elephant went extinct at least 30,000 years ago, although most of the fossils that have been discovered are at least 100,000 years old."[69]
"Palaeoloxodon (straight-tusked elephants; Figure 1), [...] appears in Eurasia around 0.75 million years ago (Ma) (Lister, 2016)."[69]
"Across its range from Western Europe to Japan, Palaeoloxodon probably comprised several species [including Palaeoloxodon namadicus] (Shoshani et al., 2007), and, based on morphological comparisons, all of them are considered to be derived from the African Palaeoloxodon (or Elephas) recki (Maglio, 1973; Saegusa and Gilbert, 2008), which was the predominant proboscidean lineage in Africa during the Pliocene and Pleistocene but went extinct around 100 thousand years ago (ka) (Owen-Smith, 2013). Straight-tusked elephants may have survived in mainland Eurasia until around 35 ka, although the youngest reliably dated remains are from the last interglacial, 115–130 ka (Stuart, 2005)."[69]
"The four straight-tusked elephants [genetically tested] did not cluster together within this mitochondrial [F-]clade [mito-genetic diversity of extant L. cyclotis], but formed two separate lineages that share a common ancestor with an extant L. cyclotis lineage 0.7–1.6 Ma (NN) and 1.5–3.0 Ma (WE) ago, respectively."[69]
Loxodonta
editLoxodonta, the generic name for the African elephants, is Greek for "oblique-sided tooth".[70]
Loxodonta africana
edit
"[M]itochondrial sequences of the F-clade have also been found in some L. africana individuals (Debruyne, 2005) despite the very substantial divergence of their nuclear genomes (Roca et al., 2005; Rohland et al., 2010), a pattern that has been attributed to mitochondrial gene flow from forest to savanna elephants (Roca et al., 2005)."[69]
The average brain size of an adult African elephant, Loxodonta africana, imaged at third down on the right, is around 4,200 gm.[14]
Loxodonta cyclotis
edit
At the left is the genetically confirmed separate species, the African forest elephant, Loxodonta cyclotis. Its brain size or weight may not as yet been estimated from living adults.
Elephas maximus
edit
The living elephant shown on the right is the Indian elephant, Elephas maximus. The image is of an adult male. Its brain weight is comparable to the African elephant.
"Should [an adult Indian elephant named] Happy be considered, in legal terms, a person? Which is to say, an entity capable of possessing at least some rights historically reserved for humans alone—beginning with a right to be free?"[71]
"In considering Happy’s circumstances and what might be done to improve them, should something more than animal-welfare laws and zoo regulations—which the Bronx Zoo has not violated, but arguably are inadequate—be invoked?"[71]
"Elephants, attest scientists who filed affidavits in Happy’s case, are highly self-aware, are emotional, make choices, and have a rich sense of both past and future. (Happy, in fact, was the star of a landmark 2006 Science study describing how elephants can recognize themselves in mirrors, which is considered a measure of especially human-like awareness.)"[71]
"Elephants share many key traits of autonomy with humans."[72]
"Respect for autonomy underlies our own legal right to physical liberty. Extending that to elephants is simply a matter of equality."[73]
"More to the philosophical point, the Wildlife Conservation Society cites rulings against similar Nonhuman Rights Project lawsuits filed on behalf of captive chimpanzees. According to those decisions, rights belong only to those who can also accept moral responsibility and social duties—which even the smartest animals can’t."[71]
"The rulings have been criticized, though, both by scientists who insisted that chimps do in fact have responsibilities within their own societies and by some legal theorists who don’t necessarily support chimp rights but fear a rationale that could threaten many human beings. The rights of an infant or an elderly grandmother with severe dementia are hardly contingent on the duties they fulfill."[71]
"Does an intelligent nonhuman animal who thinks and plans and appreciates life as human beings do have the right to the protection of the law against arbitrary cruelties and enforced detention? This is not merely a definitional question, but a deep dilemma of ethics and policy that deserves our attention."[74]
"Extending rights to animals could ultimately erode our own. Courts and society might, with this new paradigm, be tempted not only to look at more intelligent animals as being like humans, but start to think of less intelligent humans a little more like animals."[75]
"We kill millions of animals a day for food. If they have the right to bodily liberty, it’s basically a holocaust."[76]
"Expanding rights to women, racial minorities, and children didn’t erode the rights of property-holding white men, and implications for other species are immaterial to the question of elephant or chimpanzee rights. Lawsuits involving other species and other rights would certainly follow—but those deserve to be addressed case by case rather than forestalled en masse because it’s uncomfortable to consider what they imply for animals we eat."[71]
"Outside the United States, an Argentine court granted freedom to an orangutan at the Buenos Aires Zoo; courts in the Indian state of Uttarakhand ruled that animals both wild and domestic are not property but “legal entities” on whose behalf humans must act as guardians. The European Union, New Zealand, and Quebec explicitly recognize animals as sentient, though the actual impact of sentience laws has been limited. Legal rights for animals are no longer a fringe idea."[71]
"Inside the United States, judges in Alaska and Illinois have started to consider the well-being of pets, rather than mere ownership claims, in divorce-custody proceedings. Though an Oregon court rejected a high-profile lawsuit that would have allowed a horse’s advocates to sue for damages caused by criminal neglect, another court in that state ruled that animals could legally be considered victims of crimes—an implicit recognition that they’re more than just property."[71]
"The Animal Legal Defense Fund, which filed the aforementioned Oregon horse case, has also pushed for animals to be covered by the Freedom of Information Act, which, by the law’s letter, applies to individuals—not individual humans. Meanwhile, Friends of Animals, another advocacy organization, has collaborated with the legal philosopher Martha Nussbaum to develop what they call a “right to ethical consideration”: In their eyes, the Nonhuman Rights Project’s focus on autonomy sets too high a cognitive bar; rights might instead be based on simpler capacities, such as emotions and imagination."[71]
"Ethicists have even suggested property rights for wild animals threatened by development, labor rights for working animals, and the use of citizenship theory as a framework for thinking about animal rights. Domestic animals might be treated as full-blown citizens; wild animals are likened to members of other nations. Even if such ideas seem impractical, they’re valuable prompts to moral imagination. What would fair-labor law look like for a chicken?"[71]
"For the most part, there’s been an invisibility to anything but humans throughout the legal system. We have to bring the animals back in."[77]
Mammuthus imperator
edit
The imperial mammoth, Mammuthus imperator, shown at the left lived in North America about 20,000 b2k.
Mammuthus primigenius
editThe "mammoths and mastodons of present-day southwestern Ohio and northwestern Kentucky were homebodies that tended to stay in one area [...] The enamel on the animals' molars gave researchers clues as to where the mammoths and mastodons lived throughout their lives and what they ate. They discovered that mammoths ate grasses and sedges, whereas mastodons preferred leaves from trees or shrubs. Mammoths favored areas near retreating ice sheets, where grasses were plentiful, and mastodons fed near forested spaces."[78]
"I suspect that this was a pretty nice place to live, relatively speaking. [These] animals probably had what they needed to survive here year-round."[79]
"A group of scientists from the United States, Sweden, Canada, and the UK, has sequenced and analyzed the complete high-quality genomes of two woolly mammoths (Mammuthus primigenius) – one from northeastern Siberia and the other from Wrangel Island, located in the Arctic Ocean."[80]
The "genomes from specimens [were] taken from the remains of two male woolly mammoths, which lived about 40,000 years apart. One had lived in Siberia and is estimated to be 44,800 years old. The other – believed to be from one of the last surviving mammoth populations – lived roughly 4,300 years ago on Wrangel Island."[80]
“We found that the genome from one of the world’s last mammoths displayed low genetic variation and a signature consistent with inbreeding, likely due to the small number of mammoths that managed to survive on Wrangel Island during the last 5,000 years of the species’ existence.”[81]
“With a complete genome and this kind of data, we can now begin to understand what made a mammoth a mammoth – when compared to an elephant – and some of the underlying causes of their extinction which is an exceptionally difficult and complex puzzle to solve.”[82]
"Through careful analysis, the researchers determined the mammoth populations had suffered and recovered from a significant setback 250,000-300,000 years ago. However, another severe decline occurred in the final days of the Ice Age, marking the end."[80]
“The dates on these current samples suggest that when Egyptians were building pyramids, there were still mammoths living on these islands.”[82]
Mammuthus jeffersonii
edit
Mammuthus jeffersonii on the right is now a considered a synonym of Mammuthus columbi, but perhaps it is a hybrid between Mammuthus columbi and Mammuthus primigenius.
Mammuthus columbi
edit
Mammuthus meridionalis
edit
Mammuthus exilis
edit

Dwarf proboscideans are known to have lived in Indonesia, the Channel Islands of California, and several islands of the Mediterranean.[83]
Elephas celebensis of Sulawesi is believed to have descended from Elephas planifrons; Palaeoloxodon falconeri, or Elephas falconeri of Malta and Sicily was only 1 m (3 ft) and had probably evolved from the straight-tusked elephant; other descendants of the straight-tusked elephant existed in Cyprus; dwarf elephants of uncertain descent lived in Crete, Cyclades, and Dodecanese while dwarf mammoths are known to have lived in Sardinia.[83] The Columbian mammoth colonised the Channel Islands of California and evolved into the pygmy mammoth reaching a height of 1.2–1.8 m (4–6 ft) and weight 200–2,000 kg (440–4,410 lb); a population of small woolly mammoths survived on Wrangel Island, now 140 km (87 mi) north of the Siberian coast, as recently as 4,000 years ago.[83] After their discovery in 1993, they were considered dwarf mammoths.[84] This classification has been re-evaluated and since the Second International Mammoth Conference in 1999, these animals are no longer considered to be true "dwarf mammoths".[85]
Primates
editPositive "selection or accelerated evolution was found in a group of genes associated with primary microcephaly (MCPH), a developmental defect characterized by a severe reduction in brain size. Seven such loci (MCPH 1 – MCPH 7) with recessive mutations that lead to MCPH have been identified to date (reviewed in [19]). Of these, MCPH 5 (or abnormal spindle-like microcephaly associated, ASPM) is essential for normal mitotic spindle function in embryonic neuroblasts, with ASPM mutations being the most common cause of MCPH in a clinical sample of humans [17,19,20]."[12]
There "is strong evidence that some nucleotide changes in ASPM were subject to positive selection in the lineage leading to humans, consistant with a possible role of ASPM in the evolutionary enlargement of human brain size [6,17,20]."[12]
Hominins
editAlthough the complexity of a central nervous system, neural net may strongly influence the minimum brain size to enable the larger ganglia to think and conceive in the abstract, an apparent maximum size of 200 cm3, or 200 gm, is all that's needed to run any size animal regardless of its tissue variations.
Homo sapiens neanderthalensis
edit"Neanderthals (or Neandertals) are our closest extinct human relatives. [They may have been] a subspecies [Homo sapiens neanderthalensis]. Our well-known [...] fossil kin lived in Eurasia 200,000 to 30,000 years ago, in the Pleistocene Epoch. [...] they used tools, buried their dead and controlled fire, among other intelligent behaviors."[86]
"Neanderthals came to Europe some 300,000 years ago. They hunted big game with stone tools. Their territory spanned Europe and Asia. They left distinctive "Mousterian" artefacts."[87]
"Work on material from Italy seems to show human settlers pushing Neanderthals out (see maps). Mousterian tools were common there 45,000 years ago, when human-made Uluzzian material first appeared. By 44,000 years ago, humans were sharing Italy with a dwindling Neanderthal population. By 42,000 years ago, the Neanderthals were gone."[87]
"Around 39,000 years ago, a Neanderthal huddled in the back of a seaside cave at Gibraltar, safe from the hyenas, lions and leopards that might have prowled outside. Under the flickering light of a campfire, he or she used a stone tool to carefully etch what looks like a grid or a hashtag [in the image at the right] onto a natural platform of bedrock."[88]
"This was intentional — this was not somebody doodling or scratching on the surface."[89]
"Neanderthals might have behaved more like Homo sapiens than previously thought: They buried their dead, they used pigments and feathers to decorate their bodies, and they may have even organized their caves."[88]
"Art is something else — it's an indication of abstract thinking."[89]
"In Gorham's Cave, Finlayson and colleagues were surprised to find a series of deeply incised parallel and crisscrossing lines when they wiped away the dirt covering a bedrock surface. The rock had been sealed under a layer of soil that was littered with Mousterian stone tools (a style long linked to Neanderthals). Radiocarbon dating indicated that this soil layer was between 38,500 and 30,500 years old, suggesting the rock art buried underneath was created sometime before then."[88]
"Gibraltar is one of the most famous sites of Neanderthal occupation. At Gorham's Cave and its surrounding caverns, archaeologists have found evidence that Neanderthals butchered seals, roasted pigeons and plucked feathers off birds of prey. In other parts of Europe, Neanderthals lived alongside humans — and may have even interbred with them. But 40,000 years ago, the southern Iberian Peninsula was a Neanderthal stronghold."[88]
"Modern humans had not spread into the area yet."[89]
"More than 50 stone-tool incisions were needed to mimic the deepest line of the grid, and between 188 and 317 total strokes were probably needed to create the entire pattern."[88]
"It's very basic. It's very simple. It's not a Venus. It's not a bison. It's not a horse."[90]
"There is a huge difference between making three lines that any 3-year-old kid would be able to make and sculpting a Venus."[90]
"My own feeling is that if Neanderthals regularly used symbols, and given their longtime occupation throughout large parts of the Old World, we probably would have found clearer evidence by now."[91]
Scientists need "more than a few scratches — deliberate or not — to identify symbolic behavior on the part of Neanderthals."[91]
"Symbols, by definition, have meanings that are shared by a group of people, and because of that, they are often repeated. By itself, this is a unique example and without any intrinsic meaning … the question is not 'Could it be symbolic?' but rather 'Was it symbolic?' And to demonstrate that, it would be very important to have repeated examples."[91]
Neanderthals had an average brain size of 1500 cm3.[14] Another source puts brain sizes from three localities as Spy I 1,305 ml, Spy II 1,553 ml, and Djebel Ihroud I 1305 ml.[92]
Homo sapiens sapiens
edit| Human brain midsagittal cut |
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


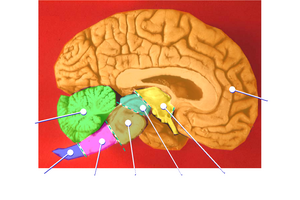
The image at the right shows the fit of a human brain into its skull with extension on the lower right to the spinal column.
The diagram at the left labels the various lobes of the human brain.
Below on the right is an animation that reveals brain lobe positioning relative to the skull.
An average brain weight for an adult human is about 1500 gms.[93] The volume varies substantially with individuals for an average of 1130 cm3 in women and 1260 cm3 in men.[94] Such a range is 1250-1450 gms.[14]
Homo erectus
edit""Peking Man," a human ancestor who lived in China between roughly 200,000 and 750,000 years ago, was a wood-working, fire-using, spear-hafting hominid ... these hominids, a form of Homo erectus, appear to have been quite meticulous about their clothing, using stone tools to soften and depress animal hides."[95]
"[S]ome of the oldest stone hand axes on Earth ... unearthed in Ethiopia ... date to 1.75 million years ago. ... fossilized H. erectus remains were also found at the same site ... These Aucheulean tools could be up to 7.8 inches (20 centimeters) long".[96]
"Javanese specimens of Homo erectus had brains about 860 cubic cm (52 cubic inches) large".[97] The Salè Homo erectus was 880 ml.[92]
Homo habilis
edit"[T]he hobbit [may have] evolved from Homo habilis, whose brains were only about 600 cubic cm (37 cubic inches)."[97]
Homo floresiensis
edit"The 18,000-year-old fossils of the extinct type of human officially known as Homo floresiensis were first discovered on the remote Indonesian island of Flores in 2003. Its squat, 3-foot-tall (1 meter) build led to the hobbit nickname."[97]
"[T]he hobbit's brain was larger than previously suggested — 426 cubic cm (nearly 26 cubic inches), instead of the commonly cited figure of 400 cubic cm. (The modern human brain is 1,300 cubic centimeters, or 79 cubic inches, large on average.)"[97]
Australopithecus afarensis
editCompared to the modern and extinct great apes, A. afarensis has reduced canines and molars, although they are still relatively larger than in modern humans. A. afarensis also has a relatively small brain size (~380–430 cm3) and a prognathic face (i.e. a face with forward projecting jaws).
Hominoids
edit{kind=link}
Rukwapithecus is "an early member of the hominoids, the group containing the great apes (gorillas, chimpanzees, bonobos, orangutans and humans) and lesser apes (gibbons)."[98]
Gorillas
edit
.png)

{kind=link}
_female_with_baby.jpg)

"In the absence of any gorilla fossil evidence, it is not possible to trace the historical genesis of these adaptations, but the three gorilla populations are assumed to have similar dietary selective regimes in comparison to other taxa that would determine the fitness of certain traits".[99]
The "whole genome of the mountain gorilla (Gorilla beringei beringei) [has been sequenced] and compared [...] with the genomes of three other Gorilla subspecies – the western lowland gorilla (G. gorilla gorilla), the Cross River gorilla (G. g. diehli), and the eastern lowland gorilla (G. b. graueri)."[100]
"They are known from two separate locations: the Virunga range of extinct volcanic mountains on the borders of the Democratic Republic of Congo, Rwanda and Uganda, and in the Bwindi Impenetrable National Park in Uganda. Some scientists believe the Bwindi gorillas may be a separate subspecies."[100]
“Mountain gorillas are among the most intensively studied primates in the wild, but this is the first in-depth, whole-genome analysis.”[100]
“Three years on from sequencing the gorilla reference genome, we can now compare the genomes of all gorilla populations, including the critically endangered mountain gorilla, and begin to understand their similarities and differences, and the genetic impact of inbreeding.”[100]
"[T]hese mountain gorillas, along with eastern lowland gorillas, were two to three times less genetically diverse than gorillas from larger groups in western regions of central Africa."[100]
"Fewer harmful loss-of-function variants were found in the mountain gorilla population than in the more numerous western gorilla populations."[100]
"[M]ountain gorillas have survived in small numbers for thousands of years."[100]
Orangutans
edit.jpg)
The Tapanuli orangutan (Pongo tapanuliensis) is a species of orangutan restricted to South Tapanuli Regency in the island of Sumatra in Indonesia.[101] It is one of three known species of orangutan, alongside the Sumatran orangutan (P. abelii), found farther northwest on the island, and Bornean orangutan (P. pygmaeus). It was described as a distinct species in 2017.[102] As of 2020, there are roughly 800 individuals of this species and it is currently on the critically endangered species list.[103]
Chimpanzees
edit
The range of brain sizes for chimpanzees is 330-430 gms.[14]
The bonobos, an individual adult is in the image at the right, are an intelligent species Pan paniscus on Earth that probably have both the necessary and sufficient mental capacity to think and conceive in the abstract. Perhaps they can also use logic or at least reasoning.
Tools
edit

Nest-building by great apes is now considered to be not just animal architecture, but as an important instance of tool use.[104]
Like the other great apes, gorillas can laugh, grieve, have "rich emotional lives", develop strong family bonds, make and use tools, and think about the past and future.[105]
All of the great apes are now known to use tools.[106]
In September 2005, a two-and-a-half-year-old gorilla in the Republic of Congo was discovered using rocks to smash open palm nuts inside a game sanctuary.[107]
In 1960, Jane Goodall observed a chimpanzee poking pieces of grass into a termite mound and then raising the grass to his mouth. After he left, Goodall approached the mound and repeated the behaviour because she was unsure what the chimpanzee was doing. She found that the termites bit onto the grass with their jaws. The chimpanzee had been using the grass as a tool to "fish" or "dip" for termites.[108]
Soon after her initial discovery of tool use, Goodall observed other chimpanzees picking up leafy twigs, stripping off the leaves and using the stems to fish for insects. This change of a leafy twig into a tool was a major discovery. Prior to this, scientists thought that only humans manufactured and used tools, and that this ability was what separated humans from other animals.[108]
Several primates have been reported as tool makers in the wild.[109]
Both bonobos and chimpanzees have been observed making "sponges" out of leaves and moss that suck up water and using these for grooming. Sumatran orangutans will take a live branch, remove twigs and leaves and sometimes the bark, before fraying or flattening the tip for use on ants or bees.[110] In the wild, mandrills have been observed to clean their ears with modified tools. Scientists filmed a large male mandrill at Chester Zoo (UK) stripping down a twig, apparently to make it narrower, and then using the modified stick to scrape dirt from underneath his toenails.[111] Captive gorillas have made a variety of tools.[112]
There are more limited reports of the closely related bonobo (Pan paniscus) using tools in the wild; it has been claimed they rarely use tools in the wild although they use tools as readily as chimpanzees when in captivity,[113] It has been reported that females of both chimpanzees and bonobos use tools more avidly than males.[114] Wild chimpanzees predominantly use tools in the context of food acquisition, while wild bonobos appear to use tools mainly for personal care (cleaning, protection from rain) and social purposes. Wild bonobos have been observed using leaves as cover for rain, or the use of branches in social displays.[113]
Researchers documented 22 occasions when wild chimpanzees on a savanna in Senegal fashioned sticks into "spears" to hunt Senegal bushbabies (lesser bushbabies) (Galago senegalensis).[115] In each case, a chimpanzee modified a branch by breaking off one or two ends and, frequently using its teeth, sharpened the stick. The tools, on average, were about 60 cm (24 in) long and 1.1 cm (0.4 in) in circumference. The chimpanzee then jabbed the spear into hollows in tree trunks where bushbabies sleep.[116]
Chimpanzees often eat the marrow of long bones of colobus monkeys with the help of small sticks, after opening the ends of the bones with their teeth.[117]
Entities
edit
John C. Lilly indicated that humans cannot have larger brains, above some size, because of insufficient neck and muscle support above some weight. Otherwise, they break their necks.
Brain-to-body mass ratio
edit
A more complex measurement, encephalization quotient, takes into account allometric effects of widely divergent body sizes across several taxa.[118][119]
Hypotheses
edit- There are, or have been, at least 23 species on Earth possessing at least as adults an abstract concept generator; i.e., the ability to think and conceive in the abstract.
- In order to conduct Original research into the possible existence of abstract concept generators within complex neural networks or systems, an understanding of universals, control groups, and at least proof of concept is needed.
See also
editReferences
edit- ↑ Teun A Van Dijk (1971). "Some problems of generative poetics". Poetics 2: 5-35. http://www.discourses.org/OldArticles/Some%20Problems%20of%20Generative%20Poetics.pdf. Retrieved 2014-07-21.
- ↑ 2.0 2.1 Speednat (17 July 2012). abstraction. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/abstraction. Retrieved 2016-05-28.
- ↑ 3.0 3.1 Speednat (29 December 2012). abstraction. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/abstraction. Retrieved 2016-05-28.
- ↑ John C. Lilly (Winter 1963). "Critical Brain Size and Language". Perspectives in Biology and Medicine 6 (2): 246-55. doi:10.1353/pbm.1963.0012. http://muse.jhu.edu/article/405958. Retrieved 2016-05-25.
- ↑ 5.0 5.1 Eclecticology (28 November 2003). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ Msh210 (3 April 2008). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ 7.0 7.1 Conrad.Irwin (10 November 2008). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ Equinox (22 February 2014). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ 64.24.209.104 (17 December 2006). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ SemperBlotto (25 January 2005). generator. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/generator. Retrieved 2 January 2019.
- ↑ SemperBlotto (25 April 2016). reification. San Francisco, California: Wikimedia Foundation, Inc. https://en.wiktionary.org/wiki/reification. Retrieved 2016-05-25.
- ↑ 12.0 12.1 12.2 12.3 12.4 12.5 12.6 12.7 Shixia Xu, Yuan Chen, Yuefeng Cheng, Dan Yang, Xuming Zhou, Junxiao Xu, Kaiya Zhou and Guang Yang (12 September 2012). "Positive selection at the ASPM gene coincides with brain size enlargements in cetaceans". Proceedings of the Royal Society B 279: 4433-4440. doi:10.1098/rspb.2012.1729. https://royalsocietypublishing.org/doi/pdf/10.1098/rspb.2012.1729. Retrieved 8 December 2021.
- ↑ WF Perrin; B Wursig; JGM Thewissen (13 November 2013). Sperm Whales (Physeter macrocephalus). NOAA. http://www.nmfs.noaa.gov/pr/species/mammals/cetaceans/spermwhale.htm. Retrieved 2014-09-23.
- ↑ 14.0 14.1 14.2 14.3 14.4 Gerhard Roth; Ursula Dicke (May 2005). "Evolution of the brain and intelligence". Trends in Cognitive Sciences 9 (5): 250-7. http://www.subjectpool.com/ed_teach/y3project/Roth2005_TICS_brain_size_and_intelligence.pdf. Retrieved 2014-09-23.
- ↑ 15.0 15.1 15.2 Ariel Mark (17 July 2018). After Thirty Years of War, the Smoke Clears. Then, the Largest Animal Ever is Found Hiding. Discovery. http://discoverytheword.com/thirty-years-war-smoke-clears-largest-animal-ever-found-hiding-ya. Retrieved 19 July 2018.
- ↑ Balaenoptera omurai. http://www.iucnredlist.org/apps/redlist/details/136623/0. Retrieved January 17, 2012.
- ↑ Wada, S.; Oishi, M.; Yamada, T.K. (2003). "A newly discovered species of living baleen whale". Nature 426 (6964): 278–281. doi:10.1038/nature02103. OCLC 110553472. PMID 14628049.
- ↑ 18.0 18.1 18.2 18.3 Raymond J. Tarpley; Sam H. Ridgway (1994). "Corpus Callosum Size in Delphinid Cetaceans". Brain Behav Evol 44: 156-65. http://www.karger.com/Article/Abstract/113587. Retrieved 2014-09-23.
- ↑ DB Tower (August 1954). "Structural and functional organization of mammalian cerebral cortex; the correlation of neurone density with brain size; cortical neurone density in the fin whale (Balaenoptera physalus L.) with a note on the cortical neurone density in the Indian elephant". J Comp Neurol 101 (1): 19-51. PMID 13211853. http://onlinelibrary.wiley.com/doi/10.1002/cne.901010103/full. Retrieved 2014-09-23.
- ↑ Kevin Spear (March 7, 2010). Killer whales: How smart are they?, In: Orlando Sentinel. http://articles.orlandosentinel.com/2010-03-06/news/os-seaworld-killer-whale-brains-20100302_1_killer-whales-orcas-dolphin-or-porpoise. Retrieved March 7, 2010.
- ↑ Lori Marino, et al. (2007). "Cetaceans Have Complex Brains for Complex Cognition". PLoS Biology 5 (e139): e139. doi:10.1371/journal.pbio.0050139. PMID 17503965. PMC 1868071. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1868071/.
- ↑ 22.0 22.1 22.2 Sarah Kaplan (3 March 2017). Caught on film for the first time: One of the world’s rarest whales. Washington, DC USA: Washington Post. https://www.washingtonpost.com/news/speaking-of-science/wp/2017/03/07/one-of-the-worlds-rarest-whales-has-been-caught-on-film-for-the-first-time/. Retrieved 2017-05-12.
- ↑ 23.0 23.1 Natacha Aguilar de Soto (3 March 2017). Caught on film for the first time: One of the world’s rarest whales. Washington, DC USA: Washington Post. https://www.washingtonpost.com/news/speaking-of-science/wp/2017/03/07/one-of-the-worlds-rarest-whales-has-been-caught-on-film-for-the-first-time/. Retrieved 2017-05-12.
- ↑ Olivier Lambert; Manuel Martínez-Cáceres; Giovanni Bianucci; Claudio Di Celma; Rodolfo Salas-Gismondi; Etienne Steurbaut; Mario Urbina; Christian de Muizon (22 May 2017). "Earliest Mysticete from the Late Eocene of Peru Sheds New Light on the Origin of Baleen Whales". Current Biology 27 (10): 1535–1541. http://www.cell.com/current-biology/fulltext/S0960-9822(17)30435-9. Retrieved 2017-06-11.
- ↑ 25.0 25.1 25.2 25.3 25.4 Tadasu K. Yamada; Shino Kitamura; Syuiti Abe; Yuko Tajima; Ayaka Matsuda; James G. Mead; Takashi F. Matsuishi (30 August 2019). "Description of a new species of beaked whale (Berardius) found in the North Pacific". Scientific Reports 9 (12723): 1-14. doi:10.1038/s41598-019-46703-w. https://www.nature.com/articles/s41598-019-46703-w.epdf?shared_access_token=VgI0k99FDGj269uA_0Qa8dRgN0jAjWel9jnR3ZoTv0PphIR8DBtoGIvwsQbJGL22qeaIthwei826TeR-e8N7I2-_HfmCNWyvOJkFzuVuPF0j2ws4rC5OBUItxJMC3kWlAEhH4TH9hu8IMcZM8G7YNA%3D%3D. Retrieved 7 September 2019.
- ↑ Kevmin (14 May 2009). "Berardiinae". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ UtherSRG (22 January 2005). "Hyperoodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ UtherSRG (22 January 2005). "Indopacetus". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ UtherSRG (22 January 2005). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ Stho002 (16 March 2014). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ Isfisk (14 August 2006). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ Isfisk (27 March 2008). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ Kempm (21 November 2006). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ Stho002 (19 December 2009). "Mesoplodon". San Francisco, California: Wikimedia Foundation, Inc. Retrieved 7 September 2019.
{{cite web}}:|author=has generic name (help) - ↑ 35.0 35.1 Caro-Beth Stewart (June 26, 2012). Building a Bigger Dolphin Brain. American Association for the Advancement of Science. http://news.sciencemag.org/2012/06/building-bigger-dolphin-brain. Retrieved 2013-11-03.
- ↑ Michael McGowen (June 26, 2012). Building a Bigger Dolphin Brain. American Association for the Advancement of Science. http://news.sciencemag.org/2012/06/building-bigger-dolphin-brain. Retrieved 2013-11-03.
- ↑ 37.0 37.1 37.2 37.3 J. Shoshani (1998). "Understanding proboscidean evolution: a formidable task". Trends in Ecology and Evolution 13 (12): 480–87. doi:10.1016/S0169-5347(98)01491-8.
- ↑ Jonathan Kingdon (2013). Mammals of Africa. Bloomsbury. p. 173. https://books.google.com/?id=B_07noCPc4kC&printsec=frontcover#v=onepage.
- ↑ Gheerbrant, E. (2009). "Paleocene emergence of elephant relatives and the rapid radiation of African ungulates". Proceedings of the National Academy of Sciences of the United States of America 106 (26): 10717–10721. doi:10.1073/pnas.0900251106. PMID 19549873. PMC 2705600. http://www.pnas.org/content/106/26/10717.full.
- ↑ Sukumar, pp. 13–16.
- ↑ 41.0 41.1 41.2 Shoshani, J.; Walter, R. C.; Abraha, M.; Berhe, S.; Tassy, P.; Sanders, W. J.; Marchant, G. H.; Libsekal, Y. et al. (2006). "A proboscidean from the late Oligocene of Eritrea, a "missing link" between early Elephantiformes and Elephantimorpha, and biogeographic implications". Proceedings of the National Academy of Sciences 103 (46): 17296–301. doi:10.1073/pnas.0603689103. PMID 17085582. PMC 1859925. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1859925/.
- ↑ 42.0 42.1 42.2 42.3 Sukumar, pp. 16–19.
- ↑ Sukumar, p. 22.
- ↑ Sukumar, pp. 24–27.
- ↑ Larramendi A (2015). "Shoulder height, body mass and shape of proboscideans" (PDF)". Acta Palaeontologica Polonica. doi:10.4202/app.00136.2014.
- ↑ Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878". In Foster, J.R.. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin. 36. New Mexico Museum of Natural History and Science. pp. 131–138.
- ↑ Hutchinson, J. R.; Delmer, C.; Miller, C. E.; Hildebrandt, T.; Pitsillides, A. A.; Boyde, A. (2011). "From flat foot to fat foot: structure, ontogeny, function, and evolution of elephant "sixth toes"". Science 334 (6063): 1699–1703. doi:10.1126/science.1211437. PMID 22194576.
- ↑ K.S. Rajgopal (2013-09-04). The dental factor in elephant evolution, In: The Hindu. http://www.thehindu.com/sci-tech/energy-and-environment/the-dental-factor-in-elephant-evolution/article5093769.ece.
- ↑ Humanity's Grassroots: How Grazing Animals Shaped Evolution. http://www.livescience.com/13550-grazing-animals-shaped-human-evolution.html.
- ↑ West, J. B. (2002). "Why doesn't the elephant have a pleural space?". Physiology 17 (2): 47–50. doi:10.1152/nips.01374.2001. PMID 11909991. http://physiologyonline.physiology.org/content/17/2/47.full.
- ↑ Larramendi, A. (2016). "Shoulder height, body mass and shape of proboscideans". Acta Palaeontologica Polonica 61. doi:10.4202/app.00136.2014. https://www.app.pan.pl/archive/published/app61/app001362014.pdf.
- ↑ William J. Sanders, 2003, chap 10, Proboscidea, in Mikael Fortelius (ed) Geology and paleontology of the Miocene Sinap Formation, Turkey, Columbia University Press, New York
- ↑ https://chasingsabretooths.wordpress.com/2017/03/23/deinotheres-for-lunch-a-sabertooths-tough-skinned-diet/
- ↑ Athanassiou, A. 2004. On a Deinotherium (Proboscidea) finding in the Neogene of Crete. Carnets de Géologie/Notebooks on Geology.
- ↑ PaleoBiology Database: Stegodon, basic info
- ↑ H. Saegusa (8 May 2006). Comparisons of Stegodon and Elephantid Abundances in the Late Pleistocene of Southern China, In: The World of Elephants. Rome: Second International Congress. pp. 345-349. https://web.archive.org/web/20060508112826/http://www.cq.rm.cnr.it/elephants2001/pdf/345_349.pdf.
- ↑ Kellogg, M.; Burkett, S.; Dennis, T. R.; Stone, G.; Gray, B. A.; McGuire, P. M.; Zori, R. T.; Stanyon, R. (2007). "Chromosome painting in the manatee supports Afrotheria and Paenungulata". Evolutionary Biology 7: 6. doi:10.1186/1471-2148-7-6. PMID 17244368. PMC 1784077. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1784077/.
- ↑ 58.0 58.1 Ozawa, T.; Hayashi, S.; Mikhelson, V. M. (1997). "Phylogenetic position of mammoth and Steller's sea cow within tethytheria demonstrated by mitochondrial DNA sequences". Journal of Molecular Evolution 44 (4): 406–13. doi:10.1007/PL00006160. PMID 9089080.
- ↑ Shoshani, J. (2005). "Order Proboscidea". Mammal Species of the World: A Taxonomic and Geographic Reference, Volume 1 (3rd ed.). Johns Hopkins University Press. pp. 90–91. ISBN 978-0-8018-8221-0. OCLC 62265494. https://www.google.com/books?id=JgAMbNSt8ikC&pg=PA91#v=onepage.
- ↑ Shoshani, pp. 38–41.
- ↑ Debruyne, R.; Barriel, V.; Tassy, P. (2003). "Mitochondrial cytochrome b of the Lyakhov mammoth (Proboscidea, Mammalia): new data and phylogenetic analyses of Elephantidae". Molecular Phylogenetics and Evolution 26 (3): 421–34. PMID 12644401. https://web.archive.org/web/20131113024755/http://regis.cubedeglace.com/wp-content/uploads/2009/02/debruyne_et_al_20031.pdf.
- ↑ Noro, M.; Masuda, R.; Dubrovo, I. A.; Yoshida, M. C.; Kato, M. (1998). "Molecular phylogenetic inference of the woolly mammoth Mammuthus primigenius, based on complete sequences of mitochondrial cytochrome b and 12S ribosomal RNA genes". Journal of Molecular Evolution 46 (3): 314–26. doi:10.1007/PL00006308. PMID 9493356.
- ↑ Rohland, N.; Malaspinas, Anna-Sapfo; Pollack, Joshua L.; Slatkin, Montgomery; Matheus, Paul; Hofreiter, Michael (2007). "Proboscidean mitogenomics: chronology and mode of elephant evolution using mastodon as outgroup". PLoS Biology 5 (8): e207. doi:10.1371/journal.pbio.0050207. PMID 17676977. PMC 1925134. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1925134/.
- ↑ Rohland, N.; Reich, D.; Mallick, S.; Meyer, M.; Green, R. E.; Georgiadis, N. J.; Roca, A. L.; Hofreiter, M. (2010). Penny, David. ed. "Genomic DNA Sequences from Mastodon and Woolly Mammoth Reveal Deep Speciation of Forest and Savanna Elephants". PLoS Biology 8 (12): e1000564. doi:10.1371/journal.pbio.1000564. PMID 21203580. PMC 3006346. //www.ncbi.nlm.nih.gov/pmc/articles/PMC3006346/.
- ↑ Roca, Alfred L.; Ishida, Yasuko; Brandt, Adam L.; Benjamin, Neal R.; Zhao, Kai; Georgiadis, Nicholas J. (2015). "Elephant Natural History: A Genomic Perspective". Annual Review of Animal Biosciences 3 (1): 139–167. doi:10.1146/annurev-animal-022114-110838. PMID 25493538.
- ↑ Gross, L. (2006). "Reading the evolutionary history of the Woolly Mammoth in its mitochondrial genome". PLoS Biology 4 (3): e74. doi:10.1371/journal.pbio.0040074. PMID 20076539. PMC 1360100. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1360100/.
- ↑ Sukumar, pp. 46–47.
- ↑ Choi, C. (2011). Woolly Mammoths Could Be Cloned Someday, Scientist Says. Live Science. http://www.livescience.com/17386-woolly-mammoth-clone.html. Retrieved 18 September 2012.
- ↑ 69.0 69.1 69.2 69.3 69.4 69.5 Matthias Meyer; Eleftheria Palkopoulou; Sina Baleka; Mathias Stiller; Kirsty E H Penkman; Kurt W Alt; Yasuko Ishida; Dietrich Mania et al. (6 June 2017). "Palaeogenomes of Eurasian straight-tusked elephants challenge the current view of elephant evolution". eLife Genomics and Evolutionary Biology 6: e25413. doi:10.7554/eLife.25413. https://elifesciences.org/articles/25413. Retrieved 2017-06-15.
- ↑ Kalb, J. E.; Mebrate, A. (1993). Fossil Elephantoids from the Hominid-Bearing Awash Group, Middle Awash Valley, Afar Depression, Ethiopia. The American Philosophical Society. pp. 52–59. ISBN 978-0-87169-831-5.
- ↑ 71.00 71.01 71.02 71.03 71.04 71.05 71.06 71.07 71.08 71.09 Brandon Keim (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Lucy Bates; Richard Byrne (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Steven Wise (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Eugene Fahey (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Richard Cupp (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Richard Epstein (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Irus Braverman (28 December 2018). An Elephant’s Personhood on Trial A legal case involving a famous solitary elephant poses a fundamental question about animals’ rights. Atlanta, Georgia USA: The Atlantic. https://www.theatlantic.com/science/archive/2018/12/happy-elephant-personhood/578818/. Retrieved 3 January 2019.
- ↑ Laura Geggel (July 28, 2014). Mammoths and Mastodons of the Ohio Valley Were Homebodies. LiveScience.com. http://www.livescience.com/47054-mammoths-mastodons-diet-roaming.html. Retrieved 2014-09-07.
- ↑ Brooke Crowley (July 28, 2014). Mammoths and Mastodons of the Ohio Valley Were Homebodies. LiveScience.com. http://www.livescience.com/47054-mammoths-mastodons-diet-roaming.html. Retrieved 2014-09-07.
- ↑ 80.0 80.1 80.2 Eleftheria Palkopoulou (23 April 2015). Genetic Scientists Sequence Complete Genomes of Two Woolly Mammoths. Sci-News. http://www.sci-news.com/genetics/science-genome-woolly-mammoth-02727.html. Retrieved 2015-06-21.
- ↑ Love Dalén (23 April 2015). Genetic Scientists Sequence Complete Genomes of Two Woolly Mammoths. Sci-News. http://www.sci-news.com/genetics/science-genome-woolly-mammoth-02727.html. Retrieved 2015-06-21.
- ↑ 82.0 82.1 Hendrik Poinar (23 April 2015). Genetic Scientists Sequence Complete Genomes of Two Woolly Mammoths. Sci-News. http://www.sci-news.com/genetics/science-genome-woolly-mammoth-02727.html. Retrieved 2015-06-21.
- ↑ 83.0 83.1 83.2 Sukumar, pp. 31–33.
- ↑ Vartanyan, S. L., Garutt, V. E., Sher, A. V.; Garutt; Sher (1993). "Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic". Nature 362 (6418): 337–40. doi:10.1038/362337a0. PMID 29633990.
- ↑ Tikhonov, A.; Agenbroad, L.; Vartanyan, S. (2003). "Comparative analysis of the mammoth populations on Wrangel Island and the Channel Islands". Deinsea 9: 415–20. ISSN 0923-9308.
- ↑ Jessie Szalay (March 19, 2013). Neanderthals: Facts About Our Extinct Human Relatives. livescience. http://www.livescience.com/28036-neanderthals-facts-about-our-extinct-human-relatives.html. Retrieved 2013-11-15.
- ↑ 87.0 87.1 Catherine Brahic (20 August 2014). Neanderthal demise traced in unprecedented detail. New Scientist. http://www.newscientist.com/article/mg22329831.500-neanderthal-demise-traced-in-unprecedented-detail.html#.U_0oRihORSU. Retrieved 2014-08-26.
- ↑ 88.0 88.1 88.2 88.3 88.4 Megan Gannon (2 September 2014). Cave Carving May Be 1st Known Example of Neanderthal Rock Art. LiveScience.com. http://www.livescience.com/47640-abstract-neanderthal-cave-engraving-discovered.html. Retrieved 2014-09-09.
- ↑ 89.0 89.1 89.2 Stewart Finlayson (2 September 2014). Cave Carving May Be 1st Known Example of Neanderthal Rock Art. LiveScience.com. http://www.livescience.com/47640-abstract-neanderthal-cave-engraving-discovered.html. Retrieved 2014-09-09.
- ↑ 90.0 90.1 Jean-Jacques Hublin (2 September 2014). Cave Carving May Be 1st Known Example of Neanderthal Rock Art. LiveScience.com. http://www.livescience.com/47640-abstract-neanderthal-cave-engraving-discovered.html. Retrieved 2014-09-09.
- ↑ 91.0 91.1 91.2 Harold Dibble (2 September 2014). Cave Carving May Be 1st Known Example of Neanderthal Rock Art. LiveScience.com. http://www.livescience.com/47640-abstract-neanderthal-cave-engraving-discovered.html. Retrieved 2014-09-09.
- ↑ 92.0 92.1 Ralph L. Holloway (1981). "Volumetric and Asymmetry Determinations on Recent Hominid Endocasts: Spy I and II, Djebel Ihroud I, and the Salè Homo erectus Specimens, With Some Notes on Neandertal Brain Size". American Journal of Physical Anthropology 55: 385-93. http://www.columbia.edu/~rlh2/Spy1%262,IrhoudEndos.AJPA1981.pdf. Retrieved 2014-09-23.
- ↑ A. Parent; M.B. Carpenter (1995). Carpenter's Human Neuroanatomy. Williams & Wilkins. ISBN 978-0-683-06752-1.
- ↑ KP Cosgrove; CM Mazure; JK Staley (2007). "Evolving knowledge of sex differences in brain structure, function, and chemistry". Biol Psychiat 62 (8): 847–55. doi:10.1016/j.biopsych.2007.03.001. PMID 17544382. PMC 2711771. //www.ncbi.nlm.nih.gov/pmc/articles/PMC2711771/.
- ↑ Owen Jarus (December 31, 2012). 'Peking Man' Was a Fashion Plate. LiveScience. http://news.yahoo.com/peking-man-fashion-plate-191530302.html;_ylt=AkD.mF_Doywb34x.rZlicY57hMgF;_ylu=X3oDMTRnczd0OHZrBG1pdANUb3BTdG9yeSBTY2llbmNlU0YgRGlub3NhdXJzRm9zc2lsc1NTRgRwa2cDNjg5YTVkNzEtODE1NC0zYjRjLTg2NzYtNGRmOWNhY2Q0NmU5BHBvcwMzBHNlYwN0b3Bfc3RvcnkEdmVyAzFkMDExYjYxLTUzN2YtMTFlMi1iN2NmLWJkNjJkNzc2NzYzOA--;_ylg=X3oDMTI3bHMxdGNwBGludGwDdXMEbGFuZwNlbi11cwRwc3RhaWQDBHBzdGNhdANzY2llbmNlfGRpbm9zYXVycy1mb3NzaWxzBHB0A3NlY3Rpb25z;_ylv=3. Retrieved 2013-01-05.
- ↑ Tia Ghose (January 28, 2013). Did Rise of Ancient Human Ancestor Lead to New Stone Tools?. Yahoo! News. http://news.yahoo.com/did-rise-ancient-human-ancestor-lead-stone-tools-202218339.html. Retrieved 2013-02-02.
- ↑ 97.0 97.1 97.2 97.3 Charles Choi (April 17, 2013). The Real 'Hobbit' Had Larger Brain Than Thought. Yahoo! News. http://news.yahoo.com/real-hobbit-had-larger-brain-thought-230545891.html. Retrieved 2013-04-23.
- ↑ Tanya Lewis (May 15, 2013). Oldest Fossils Reveal When Apes & Monkeys First Diverged. Yahoo! News. http://news.yahoo.com/oldest-fossils-reveal-apes-monkeys-first-diverged-170610634.html. Retrieved 2013-05-28.
- ↑ Alison A. Elgart (June 2010). "Dental wear, wear rate, and dental disease in the African apes". American Journal of Primatology 72 (6): 481–491. doi:10.1002/ajp.20797. http://onlinelibrary.wiley.com/doi/10.1002/ajp.20797/full. Retrieved 2017-08-19.
- ↑ 100.0 100.1 100.2 100.3 100.4 100.5 100.6 Chris Tyler-Smith and Dr Aylwyn Scally (9 April 2015). "Mountain Gorillas Reveal Their Genetic Secrets". Sci-News. Retrieved 2015-06-21.
- ↑ Nater, Alexander; Mattle-Greminger, Maja P.; Nurcahyo, Anton; Nowak, Matthew G.; de Manuel, Marc; Desai, Tariq; Groves, Colin; Pybus, Marc et al. (2 November 2017). "Morphometric, Behavioral, and Genomic Evidence for a New Orangutan Species". Current Biology 27 (22): 3487–3498. doi:10.1016/j.cub.2017.09.047. PMID 29103940. http://www.cell.com/current-biology/fulltext/S0960-9822(17)31245-9.
- ↑ Reese, April (2 November 2017). "Newly discovered orangutan species is also the most endangered". Nature 551 (7679): 151. doi:10.1038/nature.2017.22934. PMID 29120449.
- ↑ Nasution, A., et al. (2018). Declining orangutan population in the unprotected forest of Batang Toru. Tropical Life Science Research, 29(2), 77- 87.
- ↑ Marchant, Linda Frances; Nishida, Toshisada (1996). Great ape societies. Cambridge University Press. pp. 226–227. ISBN 978-0-521-55536-4. https://web.archive.org/web/20140627144319/http://books.google.com/books?id=iGfjJ4lKb1IC&pg=PA226. Retrieved 4 July 2011.
- ↑ Planet Of No Apes? Experts Warn It's Close. 10 July 2012. https://www.webcitation.org/692wzYar4?url=http://www.cbsnews.com/stories/2007/09/12/tech/main3254141.shtml. CBS News Online, 2007-09-12. Retrieved 2008-03-22.
- ↑ "First Observation of Tool Use in Wild Gorillas". PLoS Biol 3 (11): e380. 2005. doi:10.1371/journal.pbio.0030380. PMID 16187795. PMC 1236726. //www.ncbi.nlm.nih.gov/pmc/articles/PMC1236726/.
- ↑ A Tough Nut To Crack For Evolution. CBS News. 18 October 2005. https://web.archive.org/web/20061114041612/http://www.cbsnews.com/stories/2005/10/18/tech/main951800.shtml. Retrieved 18 October 2006.
- ↑ 108.0 108.1 Tool use, hunting & other discoveries. The Jane Goodall Institute. http://www.janegoodall.org/chimpanzees/tool-use-hunting-other-discoveries. Retrieved August 1, 2013.
- ↑ Boesch, Christophe; Boesch-Achermann, Hedwige (2000). The Chimpanzees of the Taï Forest: Behavioural Ecology and Evolution. Oxford University Press. p. 192. ISBN 978-01985-0508-2.
- ↑ Van Schaik, C.; Fox, E.; Sitompul, A. (1996). "Manufacture and use of tools in wild Sumatran orangutans". Naturwissenschaften 83 (4): 186–188. doi:10.1007/s001140050271.
- ↑ Gill, Victoria (July 22, 2011). Mandrill monkey makes 'pedicuring' tool. BBC. http://www.bbc.co.uk/nature/14227783. Retrieved August 11, 2013.
- ↑ Vancatova, M. (2008). Gorillas and Tools - Part I. http://www.rozhlas.cz/therevealed/comments/_zprava/488947. Retrieved August 4, 2013.
- ↑ 113.0 113.1 Bonobos. ApeTag. 2010. https://web.archive.org/web/20131102050710/https://www.clemetzoo.com/apetag/Bonobos.html. Retrieved August 3, 2013.
- ↑ Gruber, T.; Clay, Z.; Zuberbühler, K. (2010). "A comparison of bonobo and chimpanzee tool use: evidence for a female bias in the Pan lineage". Animal Behaviour 80 (6): 1023–1033. doi:10.1016/j.anbehav.2010.09.005. http://www.emory.edu/LIVING_LINKS/publications/articles/Gruber_etal_2010.pdf.
- ↑ Cognitive Archaeology and Human Evolution. Cambridge University Press. 2009. p. 66. ISBN 978-0-52176-9778.
- ↑ Raffaele, Paul (2011). Among the Great Apes: Adventures on the Trail of Our Closest Relatives. Harper. p. 109. ISBN 9780061671845.
- ↑ Shipman, Pat (2011). The Animal Connection: A New Perspective on What Makes Us Human. W. W. Norton & Company. p. 95. ISBN 9780393082227.
- ↑ Development of Intelligence. Ircamera.as.arizona.edu. http://ircamera.as.arizona.edu/NatSci102/NatSci102/lectures/intelligence.htm. Retrieved 2011-05-12.
- ↑ Cairό, O. "External measures of cognition". Front Hum Neurosci 5: 108. doi:10.3389/fnhum.2011.00108. PMID 22065955. PMC 3207484. //www.ncbi.nlm.nih.gov/pmc/articles/PMC3207484/.
External links
edit- GenomeNet KEGG database
- Google scholar Advanced Scholar Search
- Home - Gene - NCBI
- NCBI All Databases Search
- Office of Scientific & Technical Information
- PsycNET
- PubChem Public Chemical Database
Learn more about Abstract concept generators |